



Introduction
As an outcome of scientific thinking, evolutionary theories change in accordance with progress made. Here, we trace the evolution of evolutionary thought through seven different research schools that have arisen since the introduction of Darwin’s Origin of Species. These schools include Darwinism, the Modern Synthesis, Micro-, Meso-, and Macroevolution, Ecology, and Reticulate Evolution. The schools of Darwinism and the Modern Synthesis together lie at the foundation of the Neo-Darwinian paradigm. This paradigm has now expanded into the schools of Microevolution, Mesoevolution, and Macroevolution that respectively study how genes, organisms, and species evolve over time. The school of Ecology instead investigates how genes, organisms, and species interact with one another and with the physical environment in space. The Eco-Evo-Devo paradigm attempts to integrate the tenets of Ecology with those of Micro-, Meso-, and Macroevolutionary research. The Reticulate Evolution school studies non-selectionist mechanisms such as symbiosis, symbiogenesis, lateral gene transfer, infective heredity, and hybridization. This paper outlines the major research directions and points of controversies that arise between these distinct schools. It furthermore situates the Extended Evolutionary Synthesis and Third Way of Evolution along these schools. The call for an Extended Evolutionary Synthesis originated in the Mesoevolution school, while scholars active in the Third Way of Evolution movement are developing ways to recognize the important contributions made by all evolution schools.
We can depict the progress in scientific thinking on evolution by means of the Flower of Evolution (Figure 1), in which each petal represents one of seven distinct evolution schools that have evolved since the introduction of evolutionary thinking some 150 years ago.
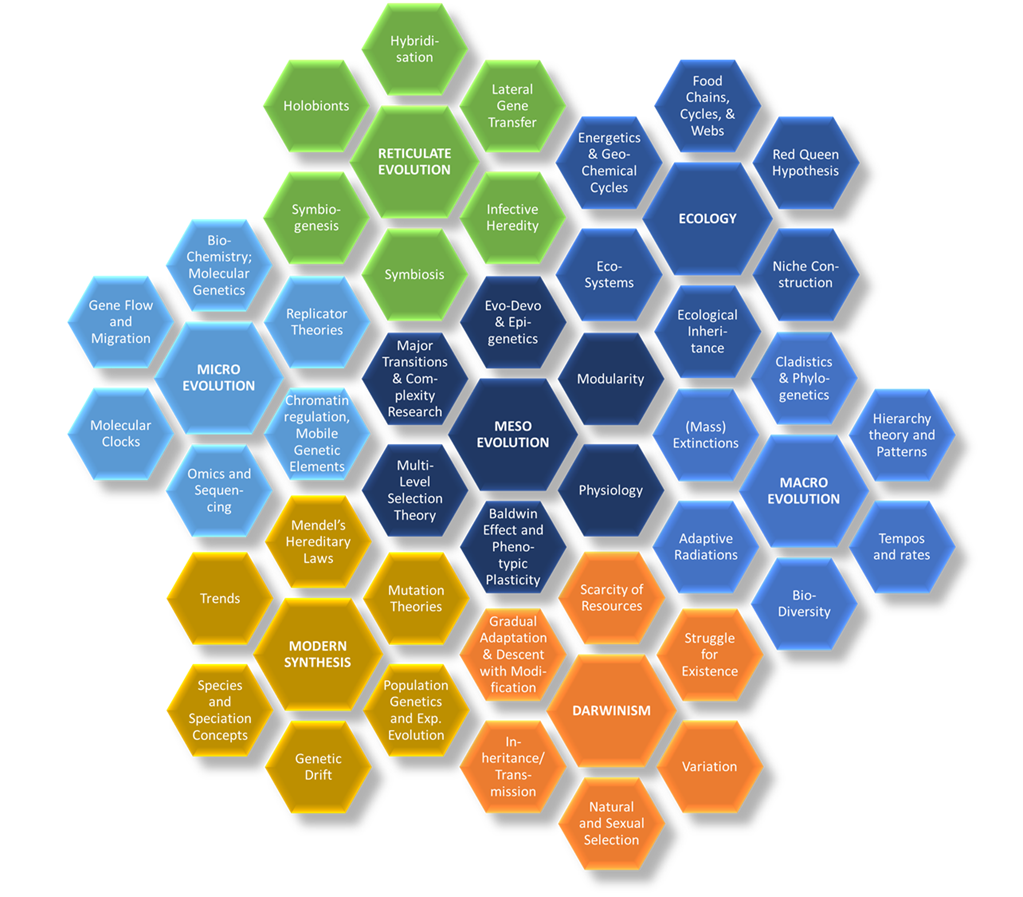
Figure 1. The Flower of Evolution. Schools in blue represent the emerging
Eco-Evo-Devo paradigm.
Several of these schools group together into larger paradigms. Darwinism and the Modern Synthesis, for example, define the Neo-Darwinian paradigm, which has long functioned as the standard paradigm within evolutionary biology. Advances made as well as attempts at meeting criticisms have resulted in an expansion of the Neo-Darwinian paradigm toward three distinct research schools: Micro-, Meso-, and Macroevolution. These examine how evolution occurs at different scales, at a micro or molecular genetic, meso or organismal, and macro or species level. But even though they focus on different phenomena, common to all three schools is that they examine evolution over time, either in deep history or during an organism’s life history.
The Ecology school traditionally endorses a spatial outlook, and focusses on how genes, organisms, and species presently interact with one another and with the abiotic environment. Today, the spatial outlook that typifies Ecology is becoming integrated with the temporal outlooks of the Micro, Meso, and Macroevolution schools into the newly emerging Ecological Evolutionary Developmental biology or Eco-Evo-Devo paradigm, depicted in Figure 1. The hope for Eco-Evo-Devo approaches is to continue expansion of the Neo-Darwinian paradigm through revision and integration.
Scholars in the Reticulate Evolution school instead call out for a more pluralistic evolutionary biology that in addition to natural selection theory recognizes the importance of non-selectionist mechanisms and processes of evolution. Darwin understood evolution by means of natural and sexual selection to result in a process of “vertical” descent with modification. Over generations through time, organisms belonging to the same species pass on their traits differentially which results in divergence that eventually underlies speciation. The reticulate evolution school instead demonstrates that evolutionary change and speciation comes about from the spatially occurring, horizontal interactions between generations of organisms belonging to different species. These mechanisms and processes include symbiosis, symbiogenesis, lateral gene transfer, hybridization, and infective heredity. Like Ecology, Reticulate Evolution thus also takes on a spatiotemporal outlook on evolution, but ecology traditionally focuses on competition between different life forms, while the Reticulate Evolution school examines all sorts of interactions, such as mutualism, commensalism, and parasitism, or the exchange of genes outside of reproduction. The different schools thus take on different perspectives and do not always agree on what evolution is. All however do provide valid data, methodologies, results, and future research directions whereby to study evolution. There currently therefore does not exist one all-encompassing theory or paradigm capable of synthesizing the different research directions. Scholars today are also questioning the need for such a paradigm, and instead recognize the pluralistic nature of evolution. Noble calls this position Biological Relativity.1
Darwinism
Although there are predecessors to evolutionary thought, Darwin’s 1859 work On the Origin of Species by Means of Natural Selection, or the Preservation of Favored Races in the Struggle for Life set in motion a new way of thinking about how species originate and change over the course of natural history. Characterizing life as a struggle for existence due to scarce environmental resources, he argued that there should naturally be a selection or favoring of organisms with traits that enhance survival. Organisms able to survive in the struggle for existence long enough to reproduce, or with traits deemed attractive to the opposite sex, will be able to pass down their traits to future generations. Organisms with maladaptive traits will not be able to pass on their traits, or will reproduce far less. The latter will naturally disappear and the former will prosper, resulting in shifts in populations and species through time that Darwin characterized as a gradual descent with modification. These ideas founded the school of Darwinism and today the concepts introduced are often called the Darwinian principles.
The Modern Synthesis
Darwin already recognized that species are related through common descent. The laws of inheritance, however, were only formulated later in time, in a non-evolutionary context, by Gregor Mendel. Mendel conjectured that trait variation is due to underlying factors, some of which are dominant and therefore expressed in the organism, others which are not, but which are nonetheless passed on recessively. Scholars later called such factors genes. A series of theoretical and experimental analyses conducted respectively by theoretical population geneticists and experimental evolutionists subsequently lead to the general conclusion that genes underlie all organismal variation. New variation was hypothesized to result from genetic mutations. Selection was considered to work on the visible organism or phenotype, where genes become expressed. In their search to explain large scale evolutionary trends, scholars would also come to recognize that selection can be intermitted by periods of no or less selection in which case gene frequencies undergo random fluctuations in number due to chance events, a process that is called drift. Selection theory thus became synthesized with Mendelian hereditary laws, gene theories, and mutation theories, and this founded the Modern Synthesis.2
Founders of the Modern Synthesis additionally introduced concepts such as gene pools, fitness landscapes, and species concepts that would provide the foundation for different perspectives wherefrom to study evolution within the Neo-Darwinian paradigm that followed. This latter paradigm evolved into three distinct research schools that study evolution at the micro (genetic), meso (organismal), and macro (species) scale. The general credo of the Neo-Darwinian paradigm, famously formulated multiple times over by Ernst Mayr, was that microevolution or small random genetic mutations suffice to explain macroevolution or the evolution of species.
Microevolution
Darwinism, the Modern Synthesis, and many tenets of the Neo-Darwinian paradigm were formulated when genes or chromosomes were still theoretical concepts. Advanced understanding of genes has come about through progress made in the fields of biochemistry and molecular genetics. Scholars can now sequence entire genomes and examine how genes flow and migrate or move within and between populations. With the help of molecular clock theories, scholars are also able to use genetic sequences to estimate relatedness and divergence in time.3
Within organisms, gene sequencing enables research on how exactly genes underlie protein synthesis, which in turn underlies tissue formation. While genes can indeed, as the founders of the Modern Synthesis thought, be implicated in the formation of anatomical form, such linkage has proven to be far from straightforward. There is hardly ever a single gene for trait x or y, for example. If traits have a genetic basis, they are brought forth by multiple genes, and the same genes are often implicated in numerous traits. The same gene, for example, can participate in the formation of the heart or the gut. A substantial part of the genome moreover is non-coding for traits and scholars are still examining the functions of these sequences. Molecular genetic research has in this regard already brought to light that genes are not the selfish replicators they once were thought to be. Genomes contain numerous mobile genetic elements that can switch location within the genome, and sometimes travel between genomes, where they possibly help in the repair of broken DNA sequences, or underlie new structural and functional changes beneficial to both the genome as well as the organism. J.A. Shapiro therefore considers genomes as “Read-Write data storage systems” rather than “Read-Only Memory information storages.”4
Mesoevolution
Much of organismal form and especially behavior cannot be reduced to genetic explanations and instead require extragenetic or epigenetic explanations as well as a more general consideration of how evolution and development interact. Stephen J. Gould was one of the first to lament that ontogeny, or research on organismal development, was long ignored by phylogenetic, evolutionary research, which focused on reconstructing evolutionary relationships of evolving species. The Weismann barrier, mentioned by other authors in this collection, lead to the assumption that what happens during an organism’s life course is unimportant for evolution because it cannot be passed on to future generations.
Such views are being revoked for several reasons. Gene-regulatory mechanisms that underlie embryogenesis and overall bodily form, for example, as well as chromosome functionality, have now been proven to undergo environmentally induced changes, and can have major evolutionary consequences. The body plan of all vertebrates, for example, is regulated by the same set of modular homeobox genes.5 Changes in the developmental expression of this homeobox gene complex have over the course of evolution resulted in variations of the different body segments that characterize the body plan of organisms as diverse as fruit flies, mice, or humans.
Beyond the genotypes from which they are constructed, organisms also demonstrate anatomical, behavioral, and cognitive plasticity and flexibility in changing environments, through their physiology or through learning. Such investigations look at how natural selection operates not only on genes but also on anatomical traits or behavior, and how multiple units and levels of selection organize hierarchically. This in turn links to more macro-oriented evolutionary research as well as to investigations into how natural selection underlies major evolutionary transitions, such as the origin of multicellular organisms, or the origin of social behavior in animals. Here too, consensus is growing that behavioral and cognitive skills, symbolization, communication, social lifeways, or culture can be subject to more inclusive or extragenetic inheritance.
Macroevolution
Macroevolution studies how evolution occurs at levels above the organism.6 Groups, populations, species, communities, or entire ecosystems can be studied for how they change over time. Macroevolutionary research can focus on how adaptive radiations and (mass) extinction events impact life’s biodiversity, or what the modes and tempos or rates of evolution are. While Darwin characterized nature as a struggle for existence, more neutral views on biodiversity have now been put forward.7 Darwin understood evolution as a process whereby species gradually evolve themselves out of existence as their lineages change into new species. He felt contrite about the lack of intermediates in the fossil record which disproved his idea. He reasoned this was due to the poor preservation of fossils in nature.
In the 1970s, Niles Eldredge and Stephen J. Gould revolutionized how we understand speciation by recognizing gaps in the fossil record not as a failure to find Darwin’s postulated intermediate forms, but as relevant data.8 They argued that if no intermediates are found, none can be postulated. Instead, some of the best preserved fossil records demonstrate that species or entire clades can sometimes appear or disappear abruptly, implying that evolution is not always a gradual process. Eldredge and Gould proposed that the fossil record is characterized by a pattern of “punctuated equilibria,” where short periods of rapid evolutionary change are interspersed by long periods of “stasis,” where no or little morphological change can be detected. Evolutionary change is thus centered around speciation events. They furthermore demonstrated that many, if not most, species evolve through splitting (cladogenesis) into sibling species rather than evolving continuously through anagenesis.
Ecology
While the micro, meso, and macroevolutionary schools study how genes, organisms, and species change through deep history or during life history events, ecological research instead examines how genes, organisms, and species behave in space, by studying how they interact with one another and with the abiotic environment. Darwin had pointed toward the abiotic and biotic environment as the locus of natural and sexual selection. The gene-centered views that followed his theories however resulted in a neglect of ecological studies that subsequently evolved outside of Modern and Neo-Darwinian Synthesis.
Ecological research on species interactions and ecosystem formation ranges from how predator and prey interact with one another and underlie food chains, cycles, and webs of life; to how co-evolution of species is shaped by the need of competing species to keep ahead of one another, as proposed by Van Valen’s “Red Queen” hypothesis (from Lewis Carroll’s Through the Looking Glass: “It takes all the running you can do, to keep in the same place”);9 to James Lovelock’s Gaia hypothesis for how life on Earth is sustained through complex biogeochemical cycles.10
Research on organism-environment relationships has furthermore made scholars realize that organisms are not merely passively selected by an active environment as Darwin or the founders of the Modern Synthesis thought. Instead, organisms can actively alter their environment though niche construction, and as such also leave an ecological footprint that can become the subject of ecological inheritance.
Reticulate Evolution
A final school that has been developing outside of the frameworks of the Modern Synthesis and the standard Neo-Darwinian paradigm is Reticulate Evolution. Reticulate evolution is evolution as it occurs by means of symbiosis, symbiogenesis, lateral gene transfer, hybridization, or infective heredity. All are inherently non-selectionist and non-Darwinian mechanisms and processes of evolution that introduce evolutionary novelty through the interaction, horizontal crossing, temporary or permanent merger of differently evolved evolutionary lineages. Evolutionary lineages thus do not so much form a tree of life but a web of life that is characterized by a reticulate or network-like structure.11
While ecological research has mainly focused on competition, symbiosis research instead has analyzed the effect of mutually beneficial, commensal (beneficial for one), and parasitic interactions on the survival and reproductive chances of the organisms engaging in symbiosis.12 The symbiotic interactions that multicellular organisms maintain with infectious agents such as microbes or viruses, for example, influence health and disease, not only during development, but also during evolution, as infections can become transmitted to future generations. Numerous genes of infectious agents have over the course of evolution even made their way into the genome of their host where they have subsequently become subject to regular inheritance. Microbiome research furthermore calls into question standard definitions of organisms as individual entities. Instead, because multicellular organisms provide an ecological life zone for numerous microorganisms, they are better characterized as holobionts.13
Holobiont formation started early on in the evolution of life. Mitochondria, for example, which are organelles found in the cytoplasm of most protist, fungi, plant, and animal cells, have a bacterial origin. Some two billion years ago, the bacteria from where they originated merged with some of the first nucleated (eukaryotic) cells and commenced a symbiosis that became permanent and hereditary. This resulted in evolution by symbiogenesis whereby the merged bacteria over time evolved into the mitochondria that typify multicellular life. The chloroplasts that characterize plant cells also evolved through symbiogenesis. Today, mitochondria and chloroplasts still contain their own DNA that is reminiscent of their bacterial origin, and this DNA is thus different from the genes found in the nucleus. Bacteria and Archaea are prokaryotes, and unlike eukaryotes, their genes are not packaged into a nucleus that is somewhat sealed off from the rest of the cell. Rather, the genes of bacteria form one long chromosome that floats freely within the cell. This enables them to rapidly exchange genes horizontally or laterally with one another in a variety of ways. Bacteria can even take up genes from dead bacteria. Such “horizontal” or “lateral” gene transfer is now well recognized to be the major means whereby bacteria acquire genes for antibiotic resistance. While horizontal gene transfer mostly occurs amongst prokaryotes, it is also known to occur between eukaryotes, and in any case, the impact of horizontal gene transfer by far extends bacteria. The rapidly developing resistance against antibiotics, for example, can affect human health and disease and thus evolution.
Hybridization is another evolutionary mechanism that underlies reticulate evolution. Long thought not to affect evolution, because some hybrids are sterile, research is demonstrating that hybridization is a means to avoid extinction, to increase genetic diversity, or to expand niche and habitat range.
On all fronts, reticulate evolution is showing that adaptation, heredity and fitness, constraints and affordances, speciation, and extinction are not merely Darwinian principles. They are also brought forth by reticulate mechanisms and processes of evolution.
Future Directions
Evolutionary thinking itself evolves in congruence with research progress. Different decades have seen various attempts at revising evolutionary thinking to either accommodate older views or to integrate new discoveries. The Modern Synthesis, for example, only emerged after decades wherein Darwinism was eclipsed over discussions on whether genetic change is continuous or discontinuous. The recognition for the need of an independent study of micro, meso, and macroevolutionary processes only emerged after the foundation of the Modern Synthesis, and such was mostly due to the rise of gene-centered views that were, in turn, made possible due to developments in molecular genetics.
Adherents of an Extended Evolutionary Synthesis have been countering gene-centered views by reappraising the role of the individual organism in evolution.14 This has first taken on the form of a synthesis between evolution and development (evo-devo). Lately, it has also focused on integrating aspects of ecology in the emerging eco-evo-devo paradigm.15 Although they have put across numerous critiques on the Neo-Darwinian paradigm, their adherents continue to seek integration with it, through revision and expansion.
Scholars of the Reticulate Evolution school have instead argued that the mechanisms and processes they study are inherently non-selectionist. At most, these mechanisms provide the evolutionary novelty that selection can work upon in a later phase of evolution. Scholars in this field have therefore called out for a new biology, that is able to better deal with the numerous ways whereby evolution occurs. Such views do not deny the importance of natural selection, they are merely asking proper recognition and independence of their specific research school.
These scholars, as well as adherents from all different evolution schools, are finding homes in the Third Way of Evolution movement that recognizes the important contributions made by the various distinctive evolution schools, with Biological Relativity being the glue that holds them together,16 The Third Way therefore does not compete with a plea for an Extended Evolutionary Synthesis, and in fact is currently fostering collaboration on important topics of teleonomy, cognition, and sociocultural evolution that as of yet remain unintegrated in general evolutionary thinking.17
At present, no school thus holds a privileged position over the other, and all have provided valid means whereby to study the pluralistic nature of evolution.
Acknowledgements
Cordial thanks to Scott Turner for his kind invitation to contribute to this inspiring feature.
Funding
Written with the financial support of the Faculdade de Ciências da Universidade de Lisboa (Faculty of Science of the University of Lisbon) and FCT, Fundação para a Ciência e a Tecnologia (the Portuguese Foundation for Science and Technology), Grant ID DL57/2016/CP1479/CT0066 and Project ID: UIDB/00678/2020.
Nathalie Gontier is is a researcher for the Faculty of Science of the University of Lisbon, and she is an Integrated Member of the Center for Philosophy of Sciences where, since 2012, she is the director of AppEEL – The Applied Evolutionary Epistemology Lab that was founded with the support of the John Templeton Foundation.
1 D. Noble, Dance to the tune of life: Biological relativity (Oxford University Press, 2017).
2 J. Huxley, Evolution: The Modern Synthesis (London: George Allen and Unwin Ltd, 1942).
3 E. Zuckerkandl, L. Pauling L, “Molecules as documents of evolutionary history,” Journal of Theoretical Biology 8, no. 2 (1965): 357–366.
4 A.J. Shapiro, Evolution: a view from the 21st century, fortified (Upper Saddle River, FT Press, 2022).
5 W. Gehring, Master control genes in development and evolution: The Homeobox story (New Haven: Yale University Press, 1998).
6 N. Eldredge, Unfinished synthesis: biological hierarchies and modern evolutionary thought (New York: Oxford University Press, 1985).
7 Robert H. MacArthur, Edward O. Wilson, The theory of island biogeography (Princeton: Princeton University Press, 1967); S. Hubbell, The unified neutral theory of biodiversity (Princeton: University of Princeton Press, 2001).
8 N, Eldredge, S. J. Gould, “Punctuated equilibria: An alternative to phyletic gradualism,” in T. M. Schopf, ed. Models in paleobiology (Freeman, Cooper & Co., 1972), pp. 82–115.
9 L.A. Van Valen, “A new evolutionary law,” Evolutionary Theory 1: 1–30.
10 J. Lovelock, Gaia: a new look at life on earth (New York: Oxford University Press, 1979).
11 W.F. Doolittle, E. Bapteste, “Pattern pluralism and the Tree of Life hypothesis,” Proc Nat Acad Sci 104, no. 7 (2007): 2043–2049.
12 J. Sapp, Evolution by association (New York: Oxford University Press, 1994).
13 R. Guerrero, L. Margulis, M. Berlanga, “Symbiogenesis: the holobiont as a unit of evolution,” Intern Microbiol 16, no. 3 (2013):133–143.
14 L. Chiu, Extended Evolutionary Synthesis: A review of the latest scientific research (John Templeton Foundation, 2022); M. Pigliucci, G. Müller, eds., Evolution: the extended synthesis (Cambridge, MA: MIT Press, 2010).
15 S.F. Gilbert, D. Epel, Ecological developmental biology (Sunderland, MA: Sinauer Associates 2008).
16 P Corning, S. Kauffman, D. Noble, J. Shapiro, R. Vane–Wright, A. Pross, Evolution on purpose: Teleonomy in living systems (Cambridge, MA: MIT Press, 2023).
17 P. A. Corning, S. Kauffman, D. Noble, J. Shapiro, R. Vane–Wright, A. Pross, Evolution on purpose: Teleonomy in living systems (Cambridge, MA: MIT Press, 2023); R.I. Vane-Wright, P.A. Corning, “Teleonomy in living systems: an overview,” Biological Journal of the Linnean Society 139, no. 4 (2023): 341–356; A. U. Igamberdiev, “Overcoming the limits of natural computation in biological evolution toward the maximization of system efficiency,” Biological Journal of the Linnean Society 139(2022): 539–554; A. Sharov, K. Kull, “Evolution and semiosis,” Bloomsbury Semiotics Volume 1: History and Semiosis (2023): 149; N. Gontier, A. Lock, C. Sinha, The Oxford handbook of human symbolic evolution (Oxford University Press, 2023).
Photo by Suzanne D. Williams on Unsplash








